De récentes études ont
mis en avant l’importance de l’épigénétique dans les mécanismes de régulation
des gènes. Parmi ces modifications, la N6-méthyladénosine (m6A) possède une
place importante puisqu’il s’agit de la modification épigénétique la plus
répandue sur les ARNm et ARNlnc des Eucaryotes (Jiang et
al., 2021). Ce changement au niveau de l’ADN peut modifier la quantité de protéines
traduite et peut avoir des conséquences diverses comme par exemple sur la
consolidation de la mémoire et de la plasticité synaptique.
La quantification de
cette marque épigénétique est principalement réalisée par des techniques basées
sur du séquençage ou de l’immunoprécipitation. Ces méthodes peuvent être
difficiles et coûteuses à mettre en place, ou alors ne permettent que d’obtenir
une résolution de l’ordre d’une centaine de nucléotides (Linder et
al., 2015). La quantification de m6A par RT-qPCR permet de mesurer le
niveau de méthylation d’une adénosine pour un coût plus faible et avec une
meilleure résolution que les techniques existantes (Castellanos-Rubio
et al., 2019).
II) Principe
Cette quantification
du niveau de méthylation des m6A est basée sur la différence
d'efficacité de deux enzymes à rétrotranscrire l'ARN en ADNc Ces deux enzymes
sont la MultiScribe™ Reverse Transcriptase (MRT) produite par Thermo Fisher
Scientific (référence # 4311235)
et la Bst DNA Polymerase, issue de Bacillus stearothermophilus, et produite
par New England Biolabs (référence #M0275). La Bst a une capacité de
rétrotranscription plus faible que l'enzyme MRT lorsque son amorce est fixée sur
un nucléotide avant un site méthylé. Cela se traduit par un ralentissement de
la vitesse de réaction de la Bst. Une plus petite quantité d'ADNc sera donc
produite par la Bst comparée à la MRT. Ce ralentissement et donc cette
différence de quantité d’ADNc pourra ainsi être quantifiée par qPCR, traduisant
le niveau de méthylation de l’adénine ciblée (Figure 1).
Figure 1 : Méthode de quantification du niveau relatif
de m6A par RT-qPCR (Figure réalisée par Elio Bobe avec Biorender)
III)
Mode opératoire
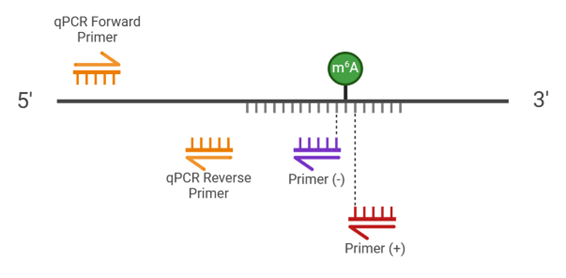
Dans un premier temps, il faut concevoir deux amorces de RT-PCR,
l'une juste avant (amorce (-)) et l'autre juste après (amorce (+)) le site
potentiellement méthylé. Ces deux amorces ne doivent pas chevaucher ce site et
en être séparées par un nucléotide (Figure 2).
L’échantillon à analyser devra subir une RT-PCR avec chacune
de ces deux amorces de manière indépendante, et ce pour chaque enzyme. Chacun
de ces échantillons devra ensuite être analysé par qPCR en utilisant deux
autres amorces situées en aval du site méthylé (Figure 2).
Figure 2 : Représentation graphique de la localisation
des amorces (Figure réalisée par Elio Bobe avec Biorender)
IV)
Présentation des résultats
Les résultats sont analysés en utilisant la formule
suivante :
Les résultats se présentent sous la forme
de graphiques :
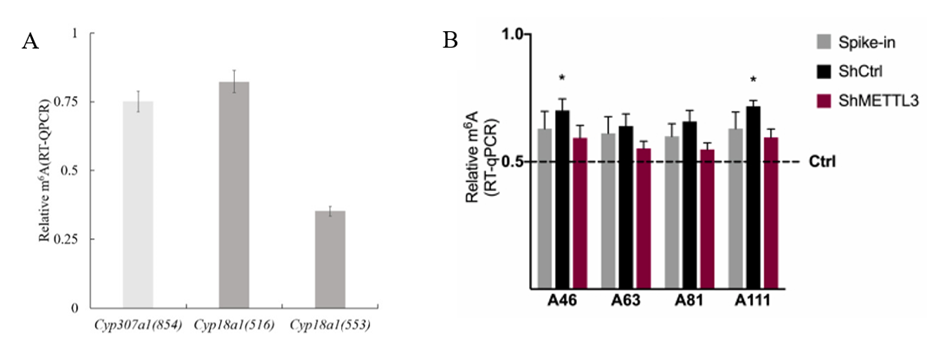
Figure 3 : Exemple de quantification de m6A
par qPCR. (A) Relative quantification of the m6A levels of
Cyp307a1 and Cyp18a1 in BmN cells (Chen et al., 2022). (B) m6A-qRT-PCR
validation of Nanopore sequencing results (Pierattini et al., 2023).
V)
Interprétation des résultats
Les résultats ayant
une valeur supérieure à 0,5 pourront être considérés comme méthylés, et ceux
ayant une valeur inférieure seront considérés comme non-méthylés. Pour comparer
des échantillons méthylés entre eux, il faudra effectuer différents tests
statistiques afin de comparer les différents niveaux relatifs de méthylation.
VI)
Intérêts et limites
Les intérêts de cette méthode sont multiples. Elle présente
un faible coût et sa facilité de mise en œuvre et d'analyse des résultats en
font un outil pratique et efficace. La précision de la méthode est également un
avantage notable, permettant une résolution d'un nucléotide. De plus, elle
offre une quantification relative du niveau de méthylation réel des ARNm.
Cependant, quelques limites sont à prendre en compte. Tout
d'abord, la nécessité de savoir quelle adénine tester implique d'avoir accès au
génome de l'échantillon. De plus, le design des amorces peut être difficile,
surtout en présence de deux sites potentiellement méthylés à proximité, ce qui
peut compliquer l'interprétation des résultats.
VII)
Références bibliographiques
Castellanos-Rubio
A, Santin I, Olazagoitia-Garmendia A, Romero-Garmendia I, Jauregi-Miguel A,
Legarda M, Bilbao JR (2019). A novel RT-QPCR-based assay for the relative
quantification of residue specific m6A RNA methylation. Sci Rep9:
4220
Chen Y, Fan B, Yasen A, Zhu J, Wang M, Shen X
(2022). YTHDF3 Is Involved in the Diapause Process of Bivoltine Bombyx mori
Strains by Regulating the Expression of Cyp307a1 and Cyp18a1 Genes in the
Ecdysone Synthesis Pathway. Biomolecules12: 1127
Jiang X, Liu B, Nie Z, Duan L, Xiong Q, Jin Z, Yang
C, Chen Y (2021). The role of m6A modification in the biological functions
and diseases. Signal Transduct Target Ther6: 74
Linder B, Grozhik AV, Olarerin-George AO, Meydan C,
Mason CE, Jaffrey SR (2015). Single-nucleotide-resolution mapping of m6A
and m6Am throughout the transcriptome. Nat Methods12: 767–772
Pierattini B, D'Agostino S, Bon C, Peruzzo O,
Alendar A, Codino A, Ros G, Persichetti F, Sanges R, Carninci P, Santoro C,
Espinoza S, Valentini P, Pandolfini L, Gustincich S (2023). SINEUP
non-coding RNA activity depends on specific N6-methyladenosine nucleotides. Mol
Ther Nucleic Acids32: 402–414